

Transdermal cannabidiol reduces inflammation and pain-related behaviours in a rat model of arthritis
D.C. Hammell,1,a L.P. Zhang,2,a F. Ma,2 S.M. Abshire,2 S.L. McIlwrath,2 A.L. Stinchcomb,1 and K.N. Westlund2
Author information ► Copyright and License information ►
The publisher's final edited version of this article is available at Eur J Pain
See other articles in PMC that cite the published article.
Abstract
Background
Current arthritis treatments often have side-effects attributable to active compounds as well as route of administration. Cannabidiol (CBD) attenuates inflammation and pain without side-effects, but CBD is hydrophobic and has poor oral bioavailability. Topical drug application avoids gastrointestinal administration, first pass metabolism, providing more constant plasma levels.
Methods
This study examined efficacy of transdermal CBD for reduction in inflammation and pain, assessing any adverse effects in a rat complete Freund’s adjuvant-induced monoarthritic knee joint model. CBD gels (0.6, 3.1, 6.2 or 62.3 mg/day) were applied for 4 consecutive days after arthritis induction. Joint circumference and immune cell invasion in histological sections were measured to indicate level of inflammation. Paw withdrawal latency (PWL) in response to noxious heat stimulation determined nociceptive sensitization, and exploratory behaviour ascertained animal’s activity level.
Results
Measurement of plasma CBD concentration provided by transdermal absorption revealed linearity with 0.6–6.2 mg/day doses. Transdermal CBD gel significantly reduced joint swelling, limb posture scores as a rating of spontaneous pain, immune cell infiltration and thickening of the synovial membrane in a dose-dependent manner. PWL recovered to near baseline level. Immunohistochemical analysis of spinal cord (CGRP, OX42) and dorsal root ganglia (TNFα) revealed dose-dependent reductions of pro-inflammatory biomarkers. Results showed 6.2 and 62 mg/day were effective doses. Exploratory behaviour was not altered by CBD indicating limited effect on higher brain function.
Conclusions
These data indicate that topical CBD application has therapeutic potential for relief of arthritis pain-related behaviours and inflammation without evident side-effects.
1. Introduction
Almost 50 million (22.2%) adult Americans over 18 were diagnosed with arthritis in 2007–2009, most prominently osteoarthritis and the autoimmune disease rheumatoid arthritis. A projected increase to 67 million is anticipated by 2030 (Centers for Disease Control and Prevention (CDC), 2010). The most effective treatment for rheumatoid arthritis is injectable fusion-proteins which sequester the most prominent proinflammatory cytokine tumour necrosis factor α (TNFα). These chimeric antibodies may halt progression of the disease, but side-effects include immune suppression (Crawford and Curtis, 2008; Furst, 2010; Hastings et al., 2010). Neurogenic drive also contributes to severity of arthritic inflammation (Sluka et al., 1994), and may contribute to its reoccurrence.
Cannabinoids and cannabinoid receptors are potential targets for reducing pain and inflammation (Clayton et al., 2002; Richardson et al., 2008; Zuardi, 2008). Cannabis sativa contains approximately 80 different cannabinoids of which Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) are primary (Mechoulam and Shvo, 1963; Mechoulam, 1970; Turner et al., 1980). These compounds are chemically similar to endogenous endocannabinoid lipid derivatives including anandamide (arachidonoylethanolamide) and 2-arachidonoylglycerol. At present THC and CBD are available combined in the oral spray Sativex® (GW Pharmaceuticals plc, Cambridge, UK) which is prescribed to adult patients for neuropathic pain. Psychoactive THC side-effects are experienced, however, and long-term use of cannabis sativa has been shown to increase risk of developing psychosis and schizophrenia (Berman et al., 2004; Malone et al., 2010). Research is ongoing to find better, effective cannabinoids and better routes of application.
CBD, while structurally similar to THC, is a non-psychoactive cannabinoid with therapeutic potential for treatment of neuropathic pain, cancer pain, multiple sclerosis and inflammation (Mechoulam and Hanus, 2002; Mechoulam et al., 2002; Burstein and Zurier, 2009). Oral bioavailability of CBD is very limited, due to first pass metabolism during digestion (Mechoulam and Hanus, 2002). The THC-like cannabinoids act at CB1 or CB2 receptors, whereas CBD-like cannabinoids have little binding affinity, leaving their role in inhibition incompletely understood. Data suggest in vitro application of CBD inhibits signalling through GPR55 and TRP channel superfamily members and in vivo oral administration is dose-dependently reducing pro-inflammatory cytokine release (Coffey et al., 1996; Malfait et al., 2000; Akopian et al., 2008; Whyte et al., 2009). More recently, CBD was successfully delivered transdermally in different species for anti-inflammatory activity (Lodzki et al., 2003; Stinchcomb et al., 2004; Paudel et al., 2010). In this study, in vivo efficacy of transdermal CBD delivery to reduce inflammation and pain-related behaviours is tested in a rat adjuvant-induced monoarthritis model with both inflammatory and neurogenic properties (Sluka et al., 1994). Nociceptive behaviour, potential adverse side-effects and inflammation-associated anatomical changes in knee joint and neuronal tissue were assessed.
2. Materials and methods
2.1 Animals
All animal procedures were approved by the University of Kentucky IACUC committee and were conducted according to guidelines for the ethical treatment of experimental animals published by the Internal Association for the Study of Pain. All experiments were conducted using 260–280 g male Sprague–Dawley rats (Harlan Laboratories, Indianapolis, IN, USA) housed in individual cages on a 12 h/12 h dark/light reversed cycle and allowed access to food and water ad libitum. A total of 54 rats were used in the experiments described here of which 21 were used as naive controls and 23 were subjected to adjuvant-induced arthritis. To induce monoarthritis, animals were anesthetized (2–4% isoflurane) and one knee joint injected with 100 μL complete Freud’s adjuvant (CFA) (DIFCO Laboratories, Detroit, MI, USA) emulsion (2 mg/mL diluted in 1:1 normal saline:peanut oil). Rats were returned to their home cages and monitored daily. Joint circumference and pain-related behaviours were assessed prior to CFA injection and daily beginning on day 3 after CFA (days 3–7).
2.2 Gel preparation
All gels, including vehicle controls, were prepared by weighing the desired amount of CBD (gift from NIDA) and dissolving it in ethanol (72.5% w/w). Once dissolved, nanopure water (Barntead NANOpure®Diamond™ ultrapure filtration system, Dubuque, IA, USA) was added followed by isopropyl myristate (Fisher Scientific, Fairlawn, NJ, USA). Carbopol® 980 polymere (Noveon Inc., Cleveland, OH, USA) was added (0.9% w/w) and the solution sonicated for 10 min to ensure complete incorporation of the Carbopol® 980. Polymerization of Carbopol ® 980 to form the hydroalcoholic gel was initiated by adding sodium hydroxide (0.1 N). Gels were then sonicated for 10 min, loaded into 1 mL syringes and sealed. Gels made just prior to the initial dosing were used for the entire week since no degradation was observed and plasma CBD concentration remained constant.
2.3 Assessment of joint inflammation
Hindlimbs were fully extended while the circumference of the knee joint was determined using a flexible tape measure wrapped around the centre of the joint. Skin temperature over the joint was measured at the patella with an infrared temperature probe (Fluke, Wilmington, North Carolina, USA). Measurements were taken on day 0, 3 and 7.
2.4 Behavioural assays
2.4.1 Spontaneous pain rating
As a measure of spontaneous pain, limb posture was scored daily in the morning while animals were in their home cages by a scientist blinded to the animal’s treatment. A subjective pain-related behavioural scale was used (Sluka et al., 1993) with 0 – normal; 1 – curling of the toes, 2 – eversion of the paw; 3 – partial weight bearing; 4 – non-weight bearing and guarding and 5 – avoidance of any contact with the hindlimb.
2.4.3 Hindpaw thermal hypersensitivity
Hindpaw heat sensitivity was assessed prior to CFA injection as well as daily starting on day 3 after inflammation, 4 h after each gel application. Paw withdrawal latency (PWL) was measured as described in our previous study (Zhang et al., 2002). Briefly, rats were acclimated for 20–30 min to individual plastic chambers (10 × 10 × 25 cm) on a glass-top table (2 mm thick). The light beam from a high-intensity projector lamp bulb (Quartzline Lamp, GE, Cleveland, OH, USA) was projected through a 5 × 10 mm aperture attached to an on/off switch with a digital timer. Maximal cut-off time for paw withdrawal reflex was set at 15 s to avoid skin damage. Both hindpaws were tested independently for 5 trials at 5 min intervals by an examiner blinded to the treatment group. The mean responses ± standard error of the mean (SEM) are reported.
2.4.4 Exploratory activity
Open-field exploratory activities were assessed in a Flexfield Animal Activity System (San Diego Instruments, San Diego, CA, USA) with Photobeam Activity System software coupled to a 486 computer (Hewlett Packard, Palo Alto, CA, USA). Animal behaviours include time spent in total exploratory activity (active time, distance travelled, total beams broken, rearing events, rearing time) and resting were recorded in a 40 × 40 × 40 cm transparent plexiglass box for 45 min prior to and at the end of the experiment (Zhang et al., 2004).
2.5 Gel administration
On day 3 after induction of monoarthritis, the back of each animal was shaved and vehicle gel or gel containing 1 or 10% CBD was applied by rubbing it into the skin for 30 s. The area of gel application and volume used corresponded to the calculated final dose of CBD, i.e. 0.62 mg CBD was dosed by applying 75 μL of 1% CBD gel on 3.5 cm2 of shaved skin of the animals’ backs, 3.1 mg CBD by giving 375 μL of 1% CBD gel on 17.5 cm2, 6.2 mg equalled 750 μL of 1% CBD gel on 35.0 cm2, and 62 mg by rubbing 750 μL of 10% CBD gel onto the skin (Table 1). Joint inflammation and nociceptive behaviour were assessed starting 4 h after gel application. On the final day of gel application after completion of all behavioural assays, rats were killed by pentobarbital overdose, blood samples were collected for CBD plasma quantification and transcardially perfused with 4% paraformaldehyde.

Table 1
Plasma concentrations (±SD) of CBD in rats with and without complete Freud’s adjuvant (CFA) induction after 4 days treatment with transdermal CBD gel.
2.6 Plasma extraction
Plasma concentration of transdermally absorbed CBD was determined. As described by Paudel et al. (2010), plasma (50 μL) was combined with 500 μL of acetonitrile (ACN) and ethyl acetate (1:1, v/v; VWR, West Chester, PA, USA), vortexed for 30 s and centrifuged at 10,000 × g for 20 min. The supernatant was transferred into a 3 mL silanized glass test tube and evaporated under nitrogen in a 37 °C water bath. It was then reconstituted in 100 μL of ACN, vortexed and sonicated for 5 min. Samples were then transferred to autosampler vials with silanized low volume inserts and 20 μL injected on the HPLC column for analysis by LC/MS.
2.7 Analytical LC/MS method
The LC/MS system used to analyse samples was comprised of a Waters Alliance 2695 pump, an autosampler, a Micromass ZQ detector and a 996 photodiode array detector with MassLynx software (Milford, MA, USA). A Symmetry® C18 column (150 × 2.1 mm, 5 μm) with a Sentry Symmetry® guard column (10 × 2.1 mm, 3.5 μm) was utilized with the LC/MS system. The ZQ detector was used with an electrospray ionization probe set for single ion monitoring for CBD quantification. Analysis was performed in negative mode for m/z 313 [CBD-H]-(dwell time: 0.3 s). Capillary and cone voltage were set at 35 kV and 40 V, respectively. Source block temperature was set at 120 °C, and desolvation temperature to 250 °C. Nitrogen was used as nebulization (flow rate: 50 l/h) and drying gas (flow rate: 450 l/h). The mobile phase was comprised of 75:25 ACN:2 mM ammonium acetate buffer w/5% ACN and used at a flow rate of 0.25 mL/min resulting in a mean CBD retention time of 5.6–5.7 min. Standard curves were linear within the range 2–300 ng/mL and concentrations of samples were determined by comparison.
2.8 Immunohistochemistry
Spinal cord, dorsal root ganglia (DRG) and knee joint synovial joint capsule membranes were excised after transcardial perfusion, post-fixed with 4% buffered paraformaldehyde, cryoprotected overnight with 30% sucrose, embedded in OCT compound (Electron Microscopy Sciences, Hatfield, PA, USA) and stored at −80 °C. Tissue was sectioned (14–20 μm) with a cryostat at −20 °C, sections collected on microscopic slides (Superfrost Plus, VWR, Radnor, PA) and stained. Synovial joint capsule membranes were stained with haematoxylin and eosin (HandE; Ricca Chemical Company, Arlington, TX, USA) and cover slipped using Permount (Fisher Scientific).
Spinal cord sections were stained using monoclonal mouse anti-OX-42 (1:1,000, CD11b/c; Abcam, Cambridge, MA, USA) and rabbit anti-CGRP (1:2000; Bachem, Torrance, CA, USA) antibodies. DRG sections were immunostained with mouse anti-TNFα (1:1000, #52B83; Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies. Primary antibodies were detected using the appropriate Alexa Fluor conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA, USA). Slides were coverslipped using Vectashield Hard Set mounting medium with DAPI (VectorLabs, Burlingame, CA, USA). Controls included the absence of staining upon omission of the primary antibody and the side to side differences between the ipsilateral and contralateral sides as internal controls.
Images of haematoxylin and eosin (H&E) staining were acquired using a Nikon Eclipse E1000 microscope equipped with a Nikon DXM 1200F digital camera and ACT-1 software. Synovial membrane thickness was measured by drawing a perpendicular line from the outer surface to the inner margin of the intimal layer of the synovial membrane in two places within each section and averaging their length (Cunnane et al., 1999).
Immunostaining of fluorescent labelled tissue was visualized using a Nikon Eclipse E1000 microscope equipped with a Cool Snap photometric Camera ES and analysed off-line with MetaMorph software. Camera settings were kept constant within an antibody staining to quantify and compare immunoreactivity. Images of the dorsal horn were analysed by outlining the substantia gelatinosa, determining and averaging the mean intensity of staining in this region. Background fluorescent intensity was measured outside of this area and subtracted. DRG sections were sampled by drawing regions of interest over five randomly selected sensory neuron somas, measuring the mean intensity and averaging for each animal sampled. Histological analysis was conducted in the four experimental groups: naive (uninjected control animals), CFA + VEH (monoarthritis treated with vehicle gel), CFA + low-dose CBD (monoarthritis treated with 0.62 or 3.1 mg/day CBD) and CFA + high-dose CBD (monoarthritis treated with 6.2 or 62.3 mg/day CBD). Mean intensities were averaged and compared across treatment groups.
2.9 Data analysis
GraphPad Prism version 6.1 for Windows (GraphPad Software, La Jolla, CA, USA) was used for all data statistical analyses. Data are presented as the mean ± standard error except where mentioned. Normally distributed data were analysed with one-way ANOVA followed by Bonferroni post hoc analysis. Pain rating score data were shown as medians and were compared using Kruskal–Wallis nonparametric analyses with Dunnett’s post hoc test. For statistical analysis of the joint circumference and behavioural results, the naive and naïve rats treated with CBD were combined for comparisons to CFA + VEH (vehicle), CFA + low-dose CBD (0.62 and 3.1 mg/day) and CFA + high-dose CBD (6.2 and 62 mg/day). Values of p ≤ 0.05 were considered significant.
3. Results
3.1 CBD plasma concentrations
Animals were treated with CBD gel in four different doses: 0.62, 3.1, 6.2 and 62.3 mg/day. After four consecutive days of treatment, plasma CBD concentrations in all rats were 3.8 ± 1.4 ng/mL (n = 9), 17.5 ± 4.4 ng/mL (n = 8), 33.3 ± 9.7 ng/mL (n = 8) and 1629.9 ± 379.0 ng/mL (n = 4), respectively (Table 1). The three lower doses displayed excellent linear pharmacokinetic correlation (slope = 1.0, R2 = 0.999). However, CBD plasma concentrations after application of the 62.3 mg/day dose did not follow the linear pharmacokinetic profile.
3.2 Knee joint inflammation
In naive animals the left and right joint circumferences were not different (mean ipsilateral = 58.0 ± 0.4 mm; mean contralateral = 57.7 ± 0.4 mm; n = 21). Three days after adjuvant induction of monoarthritis, significant swelling was noted on the side ipsilateral to the injection (ipsilateral = 73.6 ± 0.6 mm; contralateral = 55.8 ± 0.4 mm; n = 21, p < 0.001; one-way ANOVA, Bonferroni post hoc test) resulting in a 32.0 ± 1.1% circumference increase as shown in Fig. 1A. Daily applications of 6.2 mg CBD for four consecutive days significantly reduced the knee joint circumference from 72.0 ± 0.2 mm measured on day 3 after CFA injection to 65.6 ± 1.0 mm on day 7 (p < 0.05). Use of 62.3 mg/day CBD gel similarly reduced the ipsilateral joint circumference increase from 30.9 ± 0.3% to 16.7 ± 2.8% (ipsilateral circumference on day 3 = 72.0 ± 0.2 mm; day 7 = 65.6 ± 0.7 mm; p < 0.01; one-way ANOVA, Bonferroni post hoc test). Lower doses of CBD had no effect on CFA-induced oedema (Fig. 1A).
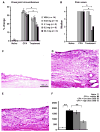
Figure 1
Assessment of knee joint inflammation. (A) Ipsilateral knee joint circumference was significantly increased in rats with adjuvant-induced monoarthritis and significantly decreased after four consecutive days of transdermal cannabidiol (CBD) treatment ...
At experiment end histological analysis after H&E staining determined that the synovial membrane was thickened 7 days after intra-articular CFA injection (Fig. 1C and D). Transdermal application of 6.2 mg/day CBD for 4 days reduced this pathological change (Fig. 1E). In naive animals the synovial membrane measured an average of 201 ± 17 μm (n = 4) from outer surface to its inner margin (Fig. 1F). This increased almost 9-fold after adjuvant-induced monoarthritis to a width of 1788 ± 128 μm (p < 0.001, one-way ANOVA, Bonferroni post hoc test n = 5). Application of the high doses of CBD (high dose = 6.2 or 62.3 mg) significantly reduced the membrane thickness by more than 50% to 767 ± 111 μm (n = 7, p < 0.001, one-way ANOVA, Bonferroni post hoc test). Treatment with the two lower doses of CBD (low dose = 0.62 or 3.1 mg) did not alter synovial membrane thickness (1631 ± 159 μm, n = 4).
3.3 Spontaneous pain rating scores
Limb posture scores as a rating of spontaneous pain were high on day 3 (median score 4) in all animals with adjuvant-induced monoarthritis. On day 7, after 4 days of transcutaneous treatment with 6.2 and 62.3 mg/day CBD gel, pain scores were significantly improved (median 1.5) compared to animals in the vehicle control group (median 3.5; p < 0.05; Kruskal–Wallis test, Dunnett’s post hoc test) (Fig. 1B). Pain scores of animals that received 0.6 and 3.1 mg/day CBD were not different from vehicle controls. All naive rats scored 0 in this test.
3.4 Secondary hindpaw heat hypersensitivity
Baseline paw withdrawal latencies were similar in all experimental animals (10–12 s). Hypersensitivity to noxious heat was detected in all animals with adjuvant-induced monoarthritis. Average paw withdrawal latency (PWL) in response to radiant heat applied to the plantar surface of the same side hindpaw, was significantly decreased on day 3 from 11.2 ± 0.2 s to 7.2 ± 0.1 s (p < 0.005, one-way ANOVA, Bonferroni post hoc test). The PWL on the contralateral side did not change (day 0 = 11.1 ± 0.2 s; day 3 = 11.1 ± 0.2 s; p > 0.05, one-way ANOVA, Bonferroni post hoc test). After 2 days of treatment with 6.2 or 62 mg/day transdermal CBD, a significant improvement of heat hypersensitivity was noted on day 5 which persisted throughout the experiment (Fig. 2A). The 6.2 mg/day doses increased PWL of the inflamed leg from 7.5 ± 0.1 s on day 3 after CFA injection to 9.4 ± 0.5 s on day 5 (p < 0.05; one-way ANOVA, Bonferroni post hoctest). The 62 mg/day increased PWL from 6.7 ± 0 ± 0.1 on day 5 (p < 0.05, one-way ANOVA, Bonferroni post hoc test). Reduction in monoarthritis-induced heat hypersensitivity was maximal after 2 days of transdermal application of 6.2 or 62 mg/day CBD, and no further improvement was evident through the remaining experimental time course. Adjuvant-induced heat hypersensitivity was not changed by transdermal application of vehicle, nor by 0.6 or 3.1 mg/day CBD (Fig. 2A). Daily application of CBD gel on naive animals did not alter heat sensitivity irrespective of the concentration used (Fig. 2B).
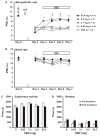
Figure 2
Transdermal cannabidiol (CBD) reduced monoarthritis-induced hind paw heat hypersensitivity. (A) Transdermal CBD significantly increased heat paw withdrawal latency in rats with monoarthritis, (B) but had no effect in naïve rats (*p < 0.01 ...
3.5 Activity level in open field
Potential for adverse side-effects on activity levels or motor abilities stemming from CBD gel application were assessed by monitoring open-field exploratory behaviour of naive animals for 45 min prior to and directly after treatment. Irrespective of the amount of CBD applied onto the back of animals, no changes were detected for total time spent in either exploratory activity or resting (Fig. 2C and D). Two other specific exploratory activities acquired in the open-field test but not affected by CBD were rearing events in which the animals rise on their hindlimbs to explore the environment and the total distance travelled during the 45 min test (data not shown). This indicates that CBD did not alter the animals’ activity levels or motor abilities.
3.6 Immunohistochemical analysis of pro-inflammatory biomarkers
Pro-inflammatory biomarkers calcitonin gene-related protein (CGRP) (Fig. 3A) and OX42 (Fig. 3B) were immunostained in the lumbar spinal cord dorsal horn. Tumour necrosis factor alpha (TNFα) (Fig. 3C) was identified in DRG. After batch staining immunohistochemical methods, the intensity of immunofluorescence was determined using computer assisted quantification. The immunoreactivity for neuropeptide CGRP was significantly increased in the superficial dorsal horn of the spinal cord in the monoarthritis group. CGRP increased from an average fluorescent intensity of 114 ± 32 (naive) to 340 ± 81 (CFA + VEH, p < 0.05, one-way ANOVA, Bonferroni post hoc test) (Fig. 4A). Treatment with high doses of CBD (6.2 and 62 mg/day) reduced immunoreactivity to the levels in naïve animals (CFA + low dose CBD = 272 ± 46, p > 0.05 vs. CFA + VEH; CFA + high-dose CBD = 144 ± 28, p < 0.05 vs. CFA + VEH; one-way ANOVA, Bonferroni post hoc test).
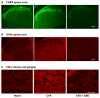
Figure 3
Immunocytochemical localization of inflammatory biomarkers. (A) CGRP and (B) OX42 are shown in ipsilateral spinal cord dorsal horn for naïve, complete Freud’s adjuvant (CFA) arthritic and cannabidiol (CBD) treated CFA arthritic rats. (C) ...
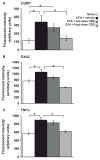
Figure 4
Quantification of pro-inflammatory biomarkers in the lumbar spinal cord and dorsal root ganglia (DRG). Increases in spinal cord (A) CGRP and (B) OX42 and (C) DRG TNFα after adjuvant-induced monoarthritis are reduced to baseline levels by high ...
Naive animals had low levels of OX42 expression in the spinal cord, a marker for activated microglia (Fig. 4B). Immunoreactivity for OX42 was significantly increased around the spinal cord central canal in lamina X in samples collected from animals 7 days after induction of monoarthritis (CFA + VEH = 1072 ± 80) compared to fluorescent intensities in naïve rats = 762 ± 38, p < 0.01, one-way ANOVA, Bonferroni post hoc test). Treatment with low doses of CBD was effective in decreasing OX42 immunoreactivity (891 ± 48, p > 0.05 vs. CFA + VEH one-way ANOVA, Bonferroni post hoc test). Treatment with high doses of CBD significantly decreased fluorescent intensity of OX42 immunoreactivity to levels below levels in the naive animals (549 ± 13, p < 0.001 vs. CFA + VEH, one-way ANOVA, Bonferroni post hoc test).
In DRG, immunoreactivity for the pro-inflammatory cytokine TNFα was significantly increased in the monoarthritic group (naive = 579 ± 33, CFA + VEH = 900 ± 63, p < 0.001; one-way ANOVA, Bonferroni post hoc test) (Fig. 4C). Treatment with high doses of CBD also reduced TNFα immunoreactivity which was equivalent to levels of naïve animals (CFA + low dose CBD = 837 ± 49, p > 0.05 vs. CFA + VEH; CFA + high-dose CBD = 620 ± 23, p < 0.001 vs. CFA + VEH; one-way ANOVA, Bonferroni post hoctest).
4. Discussion
4.1 Transdermal CBD bioactivity
Outcomes of this study indicate that topical application of CBD gel is an effective treatment for reduction in inflammation and hypersensitivity associated with the rodent adjuvant-induced monoarthritis model. Transdermal administration of CBD provided good blood absorption due to avoidance of first pass metabolism encountered by orally administered drugs. Malfait et al. (2000) saw a bell-shaped dose-dependency curve for CBD given orally or intraperitoneally to attenuate nociceptive behaviour in a collagen/CFA-induced arthritis model in mice. Similarly in this study, CBD plasma concentrations for rats dosed with 0.6, 3.1 and 6.2 mg/day exhibited an excellent linear correlation. However, the highest dose, 62.3 mg per day, did not fit into the linear pharmacokinetic profile. Dosing was increased by massaging the total amount of CBD gel into a larger skin area on the back while gel concentration (1%) remained identical. A 10% gel formulation was used for the highest transdermal CBD treatment since skin area could not be appropriately increased. The 10% gel was close to solubility saturation and may have an increased absorption rate compared to 1% formulations. The lack of increased outcome for this highest CBD concentration was potentially due to maximally activated CBD effects or capacity-limit absorption and metabolism. This would account for the flattened linear pharmacokinetic profile effect of the 62.3 mg/day dose. Spreading large quantities of gel directly on the skin over the joint itself was not feasible in this transdermal dosing paradigm and would provide opportunity for oral ingestion by the rats. Application of 10% CBD gel on skin directly overlaying an inflamed joint in patients would potentially increase local CBD concentrations and enhance effectiveness with less systemic involvement.
4.2 Efficacy of transdermal CBD
Efficacy of transdermal CBD for reduction in inflammation-associated symptoms in adjuvant-induced monoarthritic animals was determined comparing knee joint circumference and other features. The 6.2 mg/day dose optimally reduced swelling and increasing the CBD dose (62.3 mg/day) did not yield additional improvement. Likewise, increased synovial membrane thickness was reduced by the 6.2 mg/day CBD treatment. These results concur with previous studies showing orally administered CBD decreased inflammation (Malfait et al., 2000; Costa et al., 2004a,b, 2007). Decreased inflammation and reduction in secretions of pro-inflammatory and matrix-degrading effector molecules by the synovial cell connective tissue membrane lining the joints are important for symptomatic treatment of patients with rheumatoid arthritis. Pro-inflammatory and matrix-degrading effector molecules produced in excess are primary contributors to cartilage degradation over time (Ospelt et al., 2004; Neumann et al., 2010).
The improvement of pain scores provided by transdermal CBD is an indirect measure of joint inflammation and direct measure of function. The PWL in response to noxious heat stimuli was optimal with both the 6.2 and 62.3 mg/day doses. These concentrations are similar to optimal doses of intraperitoneally injected or orally administered CBD used to decrease hypersensitivity and inflammation in a collagen/CFA-induced arthritis model in mice (Malfait et al., 2000). Analogous to the results presented here, the highest dose of CBD administered in that study also did not perform as well as the next lower dose. Malfait et al. (2000) found that intraperitoneal treatments using 5 mg/kg was optimal for relief of arthritis symptoms and 25 mg/kg for orally administered treatments. Orally administered CBD at concentrations of 10–40 mg/kg were also able to inhibit thermal hypersensitivity at 3 h post carrageenan-induced inflammation of the hindpaw (Costa et al., 2004a). In the same study at 6 h post inflammation, CBD treatment with two orally administered lower doses, 5 and 7.5 mg/kg, was also effective.
4.3 Mechanisms of action
Peripheral inflammation and hypersensitivity are reversed by pharmacological inactivation of both central and peripheral neurons and central microglia (Sluka et al., 1993, 1994; Guo and Schluesener, 2006; Roberts et al., 2009). Although CBD is described as an attenuator of both mechanical and heat hypersensitivity induced by inflammatory and neuropathic pain models, the exact mechanism of action is as yet unknown (Mechoulam and Hanus, 2002; Kress and Kuner, 2009). Unlike THC and related cannabinoids, phytocannabinoid-CBD, an important bioactive component of Cannabis sativa without psychotropic effect, is an antagonist of orphan G protein-coupled receptor 55 (GPR55, a potential third metabotropic cannabinoid receptor) without binding to CB1 and CB2 receptors (Begg et al., 2005; Brown, 2007). CBD interacts with GPR55 resulting in inactivation of its pronociceptive signalling (Ryberg et al., 2007; Staton et al., 2008; Godlewski et al., 2009).
CBD also acts as an agonist for ionotropic cannabinoid receptors including chemo- and thermosensitive members of the TRP channel superfamily (TRPV1, TRPA1, TRPV2, TRPV3, TRPV4) (Fioravanti et al., 2008; Kress and Kuner, 2009). A particular focus has been on TRPA1 and TRPV1, two widely co-expressed ion channels found in CGRP expressing peptidergic nociceptors essential for neurogenic inflammation, oedema formation and inflammation-induced mechanical and thermal hypersensitivity (Davis et al., 2000; Szabo et al., 2005; Petrus et al., 2007; Kerstein et al., 2009; Akopian, 2011). Their activation by CBD in vitro results in desensitized responses following noxious stimulation with capsaicin or mustard oil, their respective agonists. This mechanism potentially decreases neuropeptide expression (Bisogno et al., 2001; Costa et al., 2004b; Akopian et al., 2008, 2009). In vivo absence or inhibition of TRPA1 results in reduced mechanical hypersensitivity in animal models of inflammation (Petrus et al., 2007; McGaraughty et al., 2010; Akopian, 2011). Absence of TRPV1 in vivo reduces inflammation-induced swelling, thermal hypersensitivity and nociceptive behaviour in various pain models (Caterina et al., 2000; Davis et al., 2000; Keeble et al., 2005; Szabo et al., 2005). In naive animals, TRPV1 immunoreactivity is localized in nociceptive primary afferents innervating the knee joint. After inflammation, TRPV1 expression increases not only in primary afferents, but is detected in synoviocytes which secrete lubricating fluid into the synovial space and function as local immune cells (Kochukov et al., 2009). Once activated, synoviocytes release pro-inflammatory cytokines and chemokines such as IL1-α, IL-6, TNFα and RANTES, as well as COX-2 (Guerne et al., 1989; Berckmans et al., 2005; Westlund et al., 2010). Released TNFα not only sensitizes primary afferents (Sorkin and Doom, 2000; Schafers and Sorkin, 2008), but also acts in an autocrine fashion, increasing the expression of TRPV1 in synoviocytes (Kochukov et al., 2009). Primary afferents are thus not only sensitized by peripheral release of pro-inflammatory cytokines, but are surrounded by cells that produce and release these molecules themselves. Release of TNFα from cultured synoviocytes from arthritic animals is reduced by application of CBD (Malfait et al., 2000) as well as by TRPV1 antagonists (Kochukov et al., 2009). Desensitization of TRPV1 by CBD could thus interrupt further progression of this cycle. Further studies are needed to identify specific receptors and mechanisms underlying the anti-inflammatory and anti-hyperalgesic effects of CBD.
The pro-inflammatory/pro-pain spiral is further driven by similar mechanisms which induce the expression of pro-inflammatory biomarkers in the DRG of the afferent nerves and centrally in the spinal cord. Transdermal CBD application was successful in decreasing monoarthritis-associated increases of pro-inflammatory biomarkers in neuronal tissues. Here, adjuvant-induced monoarthritis led to significant increases of TNFα expression in DRG which was decreased to baseline levels by transdermal CBD application.
In this study, the expression of CGRP in spinal cord was increased after peripheral inflammation as previously reported (Sluka and Westlund, 1993), and was decreased by high doses (6.2 and 62 mg/day) of transdermal CBD. Like other neuropeptides, CGRP is rapidly transported to nerve terminals for release centrally as well as peripherally where as a potent vasodilator it contributes to neurogenic inflammation (Kawasaki et al., 1988; Zhang et al., 2001). Although increases in CGRP are described in DRG after hindpaw inflammation (Nahin and Byers, 1994), no significant changes in DRG expression were observed here (data not shown), possibly due to the small number of sensory neurons innervating the joint. It is also plausible that by 7 days post monoarthritis induction, neuropeptide content has stabilized.
In monoarthritic animals, spinal cord OX42 expression is increased in activated microglia, the immune cells specific to the central nervous system, as previously described for CFA-induced ankle inflammation and trinitrobenzene sulfonic acid (TNBS)-induced pancreatitis (Shan et al., 2007; Liu et al., 2012). Treatment with high doses of transdermal CBD in this study effectively reduced OX42 expression below baseline levels, indicating reduced microglial activation. CBD also reduced microglial/macrophage spinal cord expression in a mouse encephalopathy model (Kozela et al., 2011).
4.4 Absence of psychoactive behaviours
CBD is a known non-psychoactive cannabinoid, and due to its low affinity for the CB1 receptor it would be expected that exploratory behavioural activity would be similar among treatment groups compared to negative side-effects associated with THC (Croxford, 2003; Malone et al., 2009). This was demonstrated in this study by lack of CBD-induced changes in open-field exploration among naive treatment groups. Combinatorial with psychoactivity, side-effects such as hypothermia and hypomobility induced by THC are avoided with use of CBD (Zimmer et al., 1999).
5. Conclusion
These studies demonstrate transdermal administration of CBD has long-lasting therapeutic effects without psychoactive side-effects. Thus, use of topical CBD has potential as effective treatment of arthritic symptomatology. At present, one in five (21%) adults worldwide are diagnosed with some form of arthritis by their physicians (Helmick et al., 2008). The data presented suggest transdermal CBD is a good candidate for developing improved therapies for these debilitating disease.
What’s already known about this topic?
Acknowledgments
Funding sources
This work was supported by the American Cancer Society award (RGS-00-027-04-CDD) to A.L. Stinchcomb, University of Kentucky start-up funds to K.N. Westlund and All-Tranz, Inc. Cannabidiol was a generous gift provided by NIDA.
Footnotes
Conflicts of interest
None declared.
Author contributions
D.C.H. prepared and applied the treatments, tested blood drug concentrations, performed data analysis and wrote portions of the first draft of this manuscript. L.Z. performed the drug application, behavioural tests, data analysis, statistics and produced the figures, as well as drafted portions of and edited this manuscript. F.M. performed the blinded behavioural tests, read and edited this manuscript. S.M.A. performed the blinded behavioural tests and read this manuscript. S.L.M. performed the immunostaining, data analysis, produced the figures, as well as read and edited this manuscript. A.L.S. designed the study, supplied the drug, read and edited this manuscript. K.N.W. designed the study, read and edited this manuscript. All authors discussed the results and commented on the manuscript.
References
D.C. Hammell,1,a L.P. Zhang,2,a F. Ma,2 S.M. Abshire,2 S.L. McIlwrath,2 A.L. Stinchcomb,1 and K.N. Westlund2
Author information ► Copyright and License information ►
The publisher's final edited version of this article is available at Eur J Pain
See other articles in PMC that cite the published article.
Abstract
Background
Current arthritis treatments often have side-effects attributable to active compounds as well as route of administration. Cannabidiol (CBD) attenuates inflammation and pain without side-effects, but CBD is hydrophobic and has poor oral bioavailability. Topical drug application avoids gastrointestinal administration, first pass metabolism, providing more constant plasma levels.
Methods
This study examined efficacy of transdermal CBD for reduction in inflammation and pain, assessing any adverse effects in a rat complete Freund’s adjuvant-induced monoarthritic knee joint model. CBD gels (0.6, 3.1, 6.2 or 62.3 mg/day) were applied for 4 consecutive days after arthritis induction. Joint circumference and immune cell invasion in histological sections were measured to indicate level of inflammation. Paw withdrawal latency (PWL) in response to noxious heat stimulation determined nociceptive sensitization, and exploratory behaviour ascertained animal’s activity level.
Results
Measurement of plasma CBD concentration provided by transdermal absorption revealed linearity with 0.6–6.2 mg/day doses. Transdermal CBD gel significantly reduced joint swelling, limb posture scores as a rating of spontaneous pain, immune cell infiltration and thickening of the synovial membrane in a dose-dependent manner. PWL recovered to near baseline level. Immunohistochemical analysis of spinal cord (CGRP, OX42) and dorsal root ganglia (TNFα) revealed dose-dependent reductions of pro-inflammatory biomarkers. Results showed 6.2 and 62 mg/day were effective doses. Exploratory behaviour was not altered by CBD indicating limited effect on higher brain function.
Conclusions
These data indicate that topical CBD application has therapeutic potential for relief of arthritis pain-related behaviours and inflammation without evident side-effects.
1. Introduction
Almost 50 million (22.2%) adult Americans over 18 were diagnosed with arthritis in 2007–2009, most prominently osteoarthritis and the autoimmune disease rheumatoid arthritis. A projected increase to 67 million is anticipated by 2030 (Centers for Disease Control and Prevention (CDC), 2010). The most effective treatment for rheumatoid arthritis is injectable fusion-proteins which sequester the most prominent proinflammatory cytokine tumour necrosis factor α (TNFα). These chimeric antibodies may halt progression of the disease, but side-effects include immune suppression (Crawford and Curtis, 2008; Furst, 2010; Hastings et al., 2010). Neurogenic drive also contributes to severity of arthritic inflammation (Sluka et al., 1994), and may contribute to its reoccurrence.
Cannabinoids and cannabinoid receptors are potential targets for reducing pain and inflammation (Clayton et al., 2002; Richardson et al., 2008; Zuardi, 2008). Cannabis sativa contains approximately 80 different cannabinoids of which Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) are primary (Mechoulam and Shvo, 1963; Mechoulam, 1970; Turner et al., 1980). These compounds are chemically similar to endogenous endocannabinoid lipid derivatives including anandamide (arachidonoylethanolamide) and 2-arachidonoylglycerol. At present THC and CBD are available combined in the oral spray Sativex® (GW Pharmaceuticals plc, Cambridge, UK) which is prescribed to adult patients for neuropathic pain. Psychoactive THC side-effects are experienced, however, and long-term use of cannabis sativa has been shown to increase risk of developing psychosis and schizophrenia (Berman et al., 2004; Malone et al., 2010). Research is ongoing to find better, effective cannabinoids and better routes of application.
CBD, while structurally similar to THC, is a non-psychoactive cannabinoid with therapeutic potential for treatment of neuropathic pain, cancer pain, multiple sclerosis and inflammation (Mechoulam and Hanus, 2002; Mechoulam et al., 2002; Burstein and Zurier, 2009). Oral bioavailability of CBD is very limited, due to first pass metabolism during digestion (Mechoulam and Hanus, 2002). The THC-like cannabinoids act at CB1 or CB2 receptors, whereas CBD-like cannabinoids have little binding affinity, leaving their role in inhibition incompletely understood. Data suggest in vitro application of CBD inhibits signalling through GPR55 and TRP channel superfamily members and in vivo oral administration is dose-dependently reducing pro-inflammatory cytokine release (Coffey et al., 1996; Malfait et al., 2000; Akopian et al., 2008; Whyte et al., 2009). More recently, CBD was successfully delivered transdermally in different species for anti-inflammatory activity (Lodzki et al., 2003; Stinchcomb et al., 2004; Paudel et al., 2010). In this study, in vivo efficacy of transdermal CBD delivery to reduce inflammation and pain-related behaviours is tested in a rat adjuvant-induced monoarthritis model with both inflammatory and neurogenic properties (Sluka et al., 1994). Nociceptive behaviour, potential adverse side-effects and inflammation-associated anatomical changes in knee joint and neuronal tissue were assessed.
2. Materials and methods
2.1 Animals
All animal procedures were approved by the University of Kentucky IACUC committee and were conducted according to guidelines for the ethical treatment of experimental animals published by the Internal Association for the Study of Pain. All experiments were conducted using 260–280 g male Sprague–Dawley rats (Harlan Laboratories, Indianapolis, IN, USA) housed in individual cages on a 12 h/12 h dark/light reversed cycle and allowed access to food and water ad libitum. A total of 54 rats were used in the experiments described here of which 21 were used as naive controls and 23 were subjected to adjuvant-induced arthritis. To induce monoarthritis, animals were anesthetized (2–4% isoflurane) and one knee joint injected with 100 μL complete Freud’s adjuvant (CFA) (DIFCO Laboratories, Detroit, MI, USA) emulsion (2 mg/mL diluted in 1:1 normal saline:peanut oil). Rats were returned to their home cages and monitored daily. Joint circumference and pain-related behaviours were assessed prior to CFA injection and daily beginning on day 3 after CFA (days 3–7).
2.2 Gel preparation
All gels, including vehicle controls, were prepared by weighing the desired amount of CBD (gift from NIDA) and dissolving it in ethanol (72.5% w/w). Once dissolved, nanopure water (Barntead NANOpure®Diamond™ ultrapure filtration system, Dubuque, IA, USA) was added followed by isopropyl myristate (Fisher Scientific, Fairlawn, NJ, USA). Carbopol® 980 polymere (Noveon Inc., Cleveland, OH, USA) was added (0.9% w/w) and the solution sonicated for 10 min to ensure complete incorporation of the Carbopol® 980. Polymerization of Carbopol ® 980 to form the hydroalcoholic gel was initiated by adding sodium hydroxide (0.1 N). Gels were then sonicated for 10 min, loaded into 1 mL syringes and sealed. Gels made just prior to the initial dosing were used for the entire week since no degradation was observed and plasma CBD concentration remained constant.
2.3 Assessment of joint inflammation
Hindlimbs were fully extended while the circumference of the knee joint was determined using a flexible tape measure wrapped around the centre of the joint. Skin temperature over the joint was measured at the patella with an infrared temperature probe (Fluke, Wilmington, North Carolina, USA). Measurements were taken on day 0, 3 and 7.
2.4 Behavioural assays
2.4.1 Spontaneous pain rating
As a measure of spontaneous pain, limb posture was scored daily in the morning while animals were in their home cages by a scientist blinded to the animal’s treatment. A subjective pain-related behavioural scale was used (Sluka et al., 1993) with 0 – normal; 1 – curling of the toes, 2 – eversion of the paw; 3 – partial weight bearing; 4 – non-weight bearing and guarding and 5 – avoidance of any contact with the hindlimb.
2.4.3 Hindpaw thermal hypersensitivity
Hindpaw heat sensitivity was assessed prior to CFA injection as well as daily starting on day 3 after inflammation, 4 h after each gel application. Paw withdrawal latency (PWL) was measured as described in our previous study (Zhang et al., 2002). Briefly, rats were acclimated for 20–30 min to individual plastic chambers (10 × 10 × 25 cm) on a glass-top table (2 mm thick). The light beam from a high-intensity projector lamp bulb (Quartzline Lamp, GE, Cleveland, OH, USA) was projected through a 5 × 10 mm aperture attached to an on/off switch with a digital timer. Maximal cut-off time for paw withdrawal reflex was set at 15 s to avoid skin damage. Both hindpaws were tested independently for 5 trials at 5 min intervals by an examiner blinded to the treatment group. The mean responses ± standard error of the mean (SEM) are reported.
2.4.4 Exploratory activity
Open-field exploratory activities were assessed in a Flexfield Animal Activity System (San Diego Instruments, San Diego, CA, USA) with Photobeam Activity System software coupled to a 486 computer (Hewlett Packard, Palo Alto, CA, USA). Animal behaviours include time spent in total exploratory activity (active time, distance travelled, total beams broken, rearing events, rearing time) and resting were recorded in a 40 × 40 × 40 cm transparent plexiglass box for 45 min prior to and at the end of the experiment (Zhang et al., 2004).
2.5 Gel administration
On day 3 after induction of monoarthritis, the back of each animal was shaved and vehicle gel or gel containing 1 or 10% CBD was applied by rubbing it into the skin for 30 s. The area of gel application and volume used corresponded to the calculated final dose of CBD, i.e. 0.62 mg CBD was dosed by applying 75 μL of 1% CBD gel on 3.5 cm2 of shaved skin of the animals’ backs, 3.1 mg CBD by giving 375 μL of 1% CBD gel on 17.5 cm2, 6.2 mg equalled 750 μL of 1% CBD gel on 35.0 cm2, and 62 mg by rubbing 750 μL of 10% CBD gel onto the skin (Table 1). Joint inflammation and nociceptive behaviour were assessed starting 4 h after gel application. On the final day of gel application after completion of all behavioural assays, rats were killed by pentobarbital overdose, blood samples were collected for CBD plasma quantification and transcardially perfused with 4% paraformaldehyde.
Table 1
Plasma concentrations (±SD) of CBD in rats with and without complete Freud’s adjuvant (CFA) induction after 4 days treatment with transdermal CBD gel.
2.6 Plasma extraction
Plasma concentration of transdermally absorbed CBD was determined. As described by Paudel et al. (2010), plasma (50 μL) was combined with 500 μL of acetonitrile (ACN) and ethyl acetate (1:1, v/v; VWR, West Chester, PA, USA), vortexed for 30 s and centrifuged at 10,000 × g for 20 min. The supernatant was transferred into a 3 mL silanized glass test tube and evaporated under nitrogen in a 37 °C water bath. It was then reconstituted in 100 μL of ACN, vortexed and sonicated for 5 min. Samples were then transferred to autosampler vials with silanized low volume inserts and 20 μL injected on the HPLC column for analysis by LC/MS.
2.7 Analytical LC/MS method
The LC/MS system used to analyse samples was comprised of a Waters Alliance 2695 pump, an autosampler, a Micromass ZQ detector and a 996 photodiode array detector with MassLynx software (Milford, MA, USA). A Symmetry® C18 column (150 × 2.1 mm, 5 μm) with a Sentry Symmetry® guard column (10 × 2.1 mm, 3.5 μm) was utilized with the LC/MS system. The ZQ detector was used with an electrospray ionization probe set for single ion monitoring for CBD quantification. Analysis was performed in negative mode for m/z 313 [CBD-H]-(dwell time: 0.3 s). Capillary and cone voltage were set at 35 kV and 40 V, respectively. Source block temperature was set at 120 °C, and desolvation temperature to 250 °C. Nitrogen was used as nebulization (flow rate: 50 l/h) and drying gas (flow rate: 450 l/h). The mobile phase was comprised of 75:25 ACN:2 mM ammonium acetate buffer w/5% ACN and used at a flow rate of 0.25 mL/min resulting in a mean CBD retention time of 5.6–5.7 min. Standard curves were linear within the range 2–300 ng/mL and concentrations of samples were determined by comparison.
2.8 Immunohistochemistry
Spinal cord, dorsal root ganglia (DRG) and knee joint synovial joint capsule membranes were excised after transcardial perfusion, post-fixed with 4% buffered paraformaldehyde, cryoprotected overnight with 30% sucrose, embedded in OCT compound (Electron Microscopy Sciences, Hatfield, PA, USA) and stored at −80 °C. Tissue was sectioned (14–20 μm) with a cryostat at −20 °C, sections collected on microscopic slides (Superfrost Plus, VWR, Radnor, PA) and stained. Synovial joint capsule membranes were stained with haematoxylin and eosin (HandE; Ricca Chemical Company, Arlington, TX, USA) and cover slipped using Permount (Fisher Scientific).
Spinal cord sections were stained using monoclonal mouse anti-OX-42 (1:1,000, CD11b/c; Abcam, Cambridge, MA, USA) and rabbit anti-CGRP (1:2000; Bachem, Torrance, CA, USA) antibodies. DRG sections were immunostained with mouse anti-TNFα (1:1000, #52B83; Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies. Primary antibodies were detected using the appropriate Alexa Fluor conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA, USA). Slides were coverslipped using Vectashield Hard Set mounting medium with DAPI (VectorLabs, Burlingame, CA, USA). Controls included the absence of staining upon omission of the primary antibody and the side to side differences between the ipsilateral and contralateral sides as internal controls.
Images of haematoxylin and eosin (H&E) staining were acquired using a Nikon Eclipse E1000 microscope equipped with a Nikon DXM 1200F digital camera and ACT-1 software. Synovial membrane thickness was measured by drawing a perpendicular line from the outer surface to the inner margin of the intimal layer of the synovial membrane in two places within each section and averaging their length (Cunnane et al., 1999).
Immunostaining of fluorescent labelled tissue was visualized using a Nikon Eclipse E1000 microscope equipped with a Cool Snap photometric Camera ES and analysed off-line with MetaMorph software. Camera settings were kept constant within an antibody staining to quantify and compare immunoreactivity. Images of the dorsal horn were analysed by outlining the substantia gelatinosa, determining and averaging the mean intensity of staining in this region. Background fluorescent intensity was measured outside of this area and subtracted. DRG sections were sampled by drawing regions of interest over five randomly selected sensory neuron somas, measuring the mean intensity and averaging for each animal sampled. Histological analysis was conducted in the four experimental groups: naive (uninjected control animals), CFA + VEH (monoarthritis treated with vehicle gel), CFA + low-dose CBD (monoarthritis treated with 0.62 or 3.1 mg/day CBD) and CFA + high-dose CBD (monoarthritis treated with 6.2 or 62.3 mg/day CBD). Mean intensities were averaged and compared across treatment groups.
2.9 Data analysis
GraphPad Prism version 6.1 for Windows (GraphPad Software, La Jolla, CA, USA) was used for all data statistical analyses. Data are presented as the mean ± standard error except where mentioned. Normally distributed data were analysed with one-way ANOVA followed by Bonferroni post hoc analysis. Pain rating score data were shown as medians and were compared using Kruskal–Wallis nonparametric analyses with Dunnett’s post hoc test. For statistical analysis of the joint circumference and behavioural results, the naive and naïve rats treated with CBD were combined for comparisons to CFA + VEH (vehicle), CFA + low-dose CBD (0.62 and 3.1 mg/day) and CFA + high-dose CBD (6.2 and 62 mg/day). Values of p ≤ 0.05 were considered significant.
3. Results
3.1 CBD plasma concentrations
Animals were treated with CBD gel in four different doses: 0.62, 3.1, 6.2 and 62.3 mg/day. After four consecutive days of treatment, plasma CBD concentrations in all rats were 3.8 ± 1.4 ng/mL (n = 9), 17.5 ± 4.4 ng/mL (n = 8), 33.3 ± 9.7 ng/mL (n = 8) and 1629.9 ± 379.0 ng/mL (n = 4), respectively (Table 1). The three lower doses displayed excellent linear pharmacokinetic correlation (slope = 1.0, R2 = 0.999). However, CBD plasma concentrations after application of the 62.3 mg/day dose did not follow the linear pharmacokinetic profile.
3.2 Knee joint inflammation
In naive animals the left and right joint circumferences were not different (mean ipsilateral = 58.0 ± 0.4 mm; mean contralateral = 57.7 ± 0.4 mm; n = 21). Three days after adjuvant induction of monoarthritis, significant swelling was noted on the side ipsilateral to the injection (ipsilateral = 73.6 ± 0.6 mm; contralateral = 55.8 ± 0.4 mm; n = 21, p < 0.001; one-way ANOVA, Bonferroni post hoc test) resulting in a 32.0 ± 1.1% circumference increase as shown in Fig. 1A. Daily applications of 6.2 mg CBD for four consecutive days significantly reduced the knee joint circumference from 72.0 ± 0.2 mm measured on day 3 after CFA injection to 65.6 ± 1.0 mm on day 7 (p < 0.05). Use of 62.3 mg/day CBD gel similarly reduced the ipsilateral joint circumference increase from 30.9 ± 0.3% to 16.7 ± 2.8% (ipsilateral circumference on day 3 = 72.0 ± 0.2 mm; day 7 = 65.6 ± 0.7 mm; p < 0.01; one-way ANOVA, Bonferroni post hoc test). Lower doses of CBD had no effect on CFA-induced oedema (Fig. 1A).
Figure 1
Assessment of knee joint inflammation. (A) Ipsilateral knee joint circumference was significantly increased in rats with adjuvant-induced monoarthritis and significantly decreased after four consecutive days of transdermal cannabidiol (CBD) treatment ...
At experiment end histological analysis after H&E staining determined that the synovial membrane was thickened 7 days after intra-articular CFA injection (Fig. 1C and D). Transdermal application of 6.2 mg/day CBD for 4 days reduced this pathological change (Fig. 1E). In naive animals the synovial membrane measured an average of 201 ± 17 μm (n = 4) from outer surface to its inner margin (Fig. 1F). This increased almost 9-fold after adjuvant-induced monoarthritis to a width of 1788 ± 128 μm (p < 0.001, one-way ANOVA, Bonferroni post hoc test n = 5). Application of the high doses of CBD (high dose = 6.2 or 62.3 mg) significantly reduced the membrane thickness by more than 50% to 767 ± 111 μm (n = 7, p < 0.001, one-way ANOVA, Bonferroni post hoc test). Treatment with the two lower doses of CBD (low dose = 0.62 or 3.1 mg) did not alter synovial membrane thickness (1631 ± 159 μm, n = 4).
3.3 Spontaneous pain rating scores
Limb posture scores as a rating of spontaneous pain were high on day 3 (median score 4) in all animals with adjuvant-induced monoarthritis. On day 7, after 4 days of transcutaneous treatment with 6.2 and 62.3 mg/day CBD gel, pain scores were significantly improved (median 1.5) compared to animals in the vehicle control group (median 3.5; p < 0.05; Kruskal–Wallis test, Dunnett’s post hoc test) (Fig. 1B). Pain scores of animals that received 0.6 and 3.1 mg/day CBD were not different from vehicle controls. All naive rats scored 0 in this test.
3.4 Secondary hindpaw heat hypersensitivity
Baseline paw withdrawal latencies were similar in all experimental animals (10–12 s). Hypersensitivity to noxious heat was detected in all animals with adjuvant-induced monoarthritis. Average paw withdrawal latency (PWL) in response to radiant heat applied to the plantar surface of the same side hindpaw, was significantly decreased on day 3 from 11.2 ± 0.2 s to 7.2 ± 0.1 s (p < 0.005, one-way ANOVA, Bonferroni post hoc test). The PWL on the contralateral side did not change (day 0 = 11.1 ± 0.2 s; day 3 = 11.1 ± 0.2 s; p > 0.05, one-way ANOVA, Bonferroni post hoc test). After 2 days of treatment with 6.2 or 62 mg/day transdermal CBD, a significant improvement of heat hypersensitivity was noted on day 5 which persisted throughout the experiment (Fig. 2A). The 6.2 mg/day doses increased PWL of the inflamed leg from 7.5 ± 0.1 s on day 3 after CFA injection to 9.4 ± 0.5 s on day 5 (p < 0.05; one-way ANOVA, Bonferroni post hoctest). The 62 mg/day increased PWL from 6.7 ± 0 ± 0.1 on day 5 (p < 0.05, one-way ANOVA, Bonferroni post hoc test). Reduction in monoarthritis-induced heat hypersensitivity was maximal after 2 days of transdermal application of 6.2 or 62 mg/day CBD, and no further improvement was evident through the remaining experimental time course. Adjuvant-induced heat hypersensitivity was not changed by transdermal application of vehicle, nor by 0.6 or 3.1 mg/day CBD (Fig. 2A). Daily application of CBD gel on naive animals did not alter heat sensitivity irrespective of the concentration used (Fig. 2B).
Figure 2
Transdermal cannabidiol (CBD) reduced monoarthritis-induced hind paw heat hypersensitivity. (A) Transdermal CBD significantly increased heat paw withdrawal latency in rats with monoarthritis, (B) but had no effect in naïve rats (*p < 0.01 ...
3.5 Activity level in open field
Potential for adverse side-effects on activity levels or motor abilities stemming from CBD gel application were assessed by monitoring open-field exploratory behaviour of naive animals for 45 min prior to and directly after treatment. Irrespective of the amount of CBD applied onto the back of animals, no changes were detected for total time spent in either exploratory activity or resting (Fig. 2C and D). Two other specific exploratory activities acquired in the open-field test but not affected by CBD were rearing events in which the animals rise on their hindlimbs to explore the environment and the total distance travelled during the 45 min test (data not shown). This indicates that CBD did not alter the animals’ activity levels or motor abilities.
3.6 Immunohistochemical analysis of pro-inflammatory biomarkers
Pro-inflammatory biomarkers calcitonin gene-related protein (CGRP) (Fig. 3A) and OX42 (Fig. 3B) were immunostained in the lumbar spinal cord dorsal horn. Tumour necrosis factor alpha (TNFα) (Fig. 3C) was identified in DRG. After batch staining immunohistochemical methods, the intensity of immunofluorescence was determined using computer assisted quantification. The immunoreactivity for neuropeptide CGRP was significantly increased in the superficial dorsal horn of the spinal cord in the monoarthritis group. CGRP increased from an average fluorescent intensity of 114 ± 32 (naive) to 340 ± 81 (CFA + VEH, p < 0.05, one-way ANOVA, Bonferroni post hoc test) (Fig. 4A). Treatment with high doses of CBD (6.2 and 62 mg/day) reduced immunoreactivity to the levels in naïve animals (CFA + low dose CBD = 272 ± 46, p > 0.05 vs. CFA + VEH; CFA + high-dose CBD = 144 ± 28, p < 0.05 vs. CFA + VEH; one-way ANOVA, Bonferroni post hoc test).
Figure 3
Immunocytochemical localization of inflammatory biomarkers. (A) CGRP and (B) OX42 are shown in ipsilateral spinal cord dorsal horn for naïve, complete Freud’s adjuvant (CFA) arthritic and cannabidiol (CBD) treated CFA arthritic rats. (C) ...
Figure 4
Quantification of pro-inflammatory biomarkers in the lumbar spinal cord and dorsal root ganglia (DRG). Increases in spinal cord (A) CGRP and (B) OX42 and (C) DRG TNFα after adjuvant-induced monoarthritis are reduced to baseline levels by high ...
Naive animals had low levels of OX42 expression in the spinal cord, a marker for activated microglia (Fig. 4B). Immunoreactivity for OX42 was significantly increased around the spinal cord central canal in lamina X in samples collected from animals 7 days after induction of monoarthritis (CFA + VEH = 1072 ± 80) compared to fluorescent intensities in naïve rats = 762 ± 38, p < 0.01, one-way ANOVA, Bonferroni post hoc test). Treatment with low doses of CBD was effective in decreasing OX42 immunoreactivity (891 ± 48, p > 0.05 vs. CFA + VEH one-way ANOVA, Bonferroni post hoc test). Treatment with high doses of CBD significantly decreased fluorescent intensity of OX42 immunoreactivity to levels below levels in the naive animals (549 ± 13, p < 0.001 vs. CFA + VEH, one-way ANOVA, Bonferroni post hoc test).
In DRG, immunoreactivity for the pro-inflammatory cytokine TNFα was significantly increased in the monoarthritic group (naive = 579 ± 33, CFA + VEH = 900 ± 63, p < 0.001; one-way ANOVA, Bonferroni post hoc test) (Fig. 4C). Treatment with high doses of CBD also reduced TNFα immunoreactivity which was equivalent to levels of naïve animals (CFA + low dose CBD = 837 ± 49, p > 0.05 vs. CFA + VEH; CFA + high-dose CBD = 620 ± 23, p < 0.001 vs. CFA + VEH; one-way ANOVA, Bonferroni post hoctest).
4. Discussion
4.1 Transdermal CBD bioactivity
Outcomes of this study indicate that topical application of CBD gel is an effective treatment for reduction in inflammation and hypersensitivity associated with the rodent adjuvant-induced monoarthritis model. Transdermal administration of CBD provided good blood absorption due to avoidance of first pass metabolism encountered by orally administered drugs. Malfait et al. (2000) saw a bell-shaped dose-dependency curve for CBD given orally or intraperitoneally to attenuate nociceptive behaviour in a collagen/CFA-induced arthritis model in mice. Similarly in this study, CBD plasma concentrations for rats dosed with 0.6, 3.1 and 6.2 mg/day exhibited an excellent linear correlation. However, the highest dose, 62.3 mg per day, did not fit into the linear pharmacokinetic profile. Dosing was increased by massaging the total amount of CBD gel into a larger skin area on the back while gel concentration (1%) remained identical. A 10% gel formulation was used for the highest transdermal CBD treatment since skin area could not be appropriately increased. The 10% gel was close to solubility saturation and may have an increased absorption rate compared to 1% formulations. The lack of increased outcome for this highest CBD concentration was potentially due to maximally activated CBD effects or capacity-limit absorption and metabolism. This would account for the flattened linear pharmacokinetic profile effect of the 62.3 mg/day dose. Spreading large quantities of gel directly on the skin over the joint itself was not feasible in this transdermal dosing paradigm and would provide opportunity for oral ingestion by the rats. Application of 10% CBD gel on skin directly overlaying an inflamed joint in patients would potentially increase local CBD concentrations and enhance effectiveness with less systemic involvement.
4.2 Efficacy of transdermal CBD
Efficacy of transdermal CBD for reduction in inflammation-associated symptoms in adjuvant-induced monoarthritic animals was determined comparing knee joint circumference and other features. The 6.2 mg/day dose optimally reduced swelling and increasing the CBD dose (62.3 mg/day) did not yield additional improvement. Likewise, increased synovial membrane thickness was reduced by the 6.2 mg/day CBD treatment. These results concur with previous studies showing orally administered CBD decreased inflammation (Malfait et al., 2000; Costa et al., 2004a,b, 2007). Decreased inflammation and reduction in secretions of pro-inflammatory and matrix-degrading effector molecules by the synovial cell connective tissue membrane lining the joints are important for symptomatic treatment of patients with rheumatoid arthritis. Pro-inflammatory and matrix-degrading effector molecules produced in excess are primary contributors to cartilage degradation over time (Ospelt et al., 2004; Neumann et al., 2010).
The improvement of pain scores provided by transdermal CBD is an indirect measure of joint inflammation and direct measure of function. The PWL in response to noxious heat stimuli was optimal with both the 6.2 and 62.3 mg/day doses. These concentrations are similar to optimal doses of intraperitoneally injected or orally administered CBD used to decrease hypersensitivity and inflammation in a collagen/CFA-induced arthritis model in mice (Malfait et al., 2000). Analogous to the results presented here, the highest dose of CBD administered in that study also did not perform as well as the next lower dose. Malfait et al. (2000) found that intraperitoneal treatments using 5 mg/kg was optimal for relief of arthritis symptoms and 25 mg/kg for orally administered treatments. Orally administered CBD at concentrations of 10–40 mg/kg were also able to inhibit thermal hypersensitivity at 3 h post carrageenan-induced inflammation of the hindpaw (Costa et al., 2004a). In the same study at 6 h post inflammation, CBD treatment with two orally administered lower doses, 5 and 7.5 mg/kg, was also effective.
4.3 Mechanisms of action
Peripheral inflammation and hypersensitivity are reversed by pharmacological inactivation of both central and peripheral neurons and central microglia (Sluka et al., 1993, 1994; Guo and Schluesener, 2006; Roberts et al., 2009). Although CBD is described as an attenuator of both mechanical and heat hypersensitivity induced by inflammatory and neuropathic pain models, the exact mechanism of action is as yet unknown (Mechoulam and Hanus, 2002; Kress and Kuner, 2009). Unlike THC and related cannabinoids, phytocannabinoid-CBD, an important bioactive component of Cannabis sativa without psychotropic effect, is an antagonist of orphan G protein-coupled receptor 55 (GPR55, a potential third metabotropic cannabinoid receptor) without binding to CB1 and CB2 receptors (Begg et al., 2005; Brown, 2007). CBD interacts with GPR55 resulting in inactivation of its pronociceptive signalling (Ryberg et al., 2007; Staton et al., 2008; Godlewski et al., 2009).
CBD also acts as an agonist for ionotropic cannabinoid receptors including chemo- and thermosensitive members of the TRP channel superfamily (TRPV1, TRPA1, TRPV2, TRPV3, TRPV4) (Fioravanti et al., 2008; Kress and Kuner, 2009). A particular focus has been on TRPA1 and TRPV1, two widely co-expressed ion channels found in CGRP expressing peptidergic nociceptors essential for neurogenic inflammation, oedema formation and inflammation-induced mechanical and thermal hypersensitivity (Davis et al., 2000; Szabo et al., 2005; Petrus et al., 2007; Kerstein et al., 2009; Akopian, 2011). Their activation by CBD in vitro results in desensitized responses following noxious stimulation with capsaicin or mustard oil, their respective agonists. This mechanism potentially decreases neuropeptide expression (Bisogno et al., 2001; Costa et al., 2004b; Akopian et al., 2008, 2009). In vivo absence or inhibition of TRPA1 results in reduced mechanical hypersensitivity in animal models of inflammation (Petrus et al., 2007; McGaraughty et al., 2010; Akopian, 2011). Absence of TRPV1 in vivo reduces inflammation-induced swelling, thermal hypersensitivity and nociceptive behaviour in various pain models (Caterina et al., 2000; Davis et al., 2000; Keeble et al., 2005; Szabo et al., 2005). In naive animals, TRPV1 immunoreactivity is localized in nociceptive primary afferents innervating the knee joint. After inflammation, TRPV1 expression increases not only in primary afferents, but is detected in synoviocytes which secrete lubricating fluid into the synovial space and function as local immune cells (Kochukov et al., 2009). Once activated, synoviocytes release pro-inflammatory cytokines and chemokines such as IL1-α, IL-6, TNFα and RANTES, as well as COX-2 (Guerne et al., 1989; Berckmans et al., 2005; Westlund et al., 2010). Released TNFα not only sensitizes primary afferents (Sorkin and Doom, 2000; Schafers and Sorkin, 2008), but also acts in an autocrine fashion, increasing the expression of TRPV1 in synoviocytes (Kochukov et al., 2009). Primary afferents are thus not only sensitized by peripheral release of pro-inflammatory cytokines, but are surrounded by cells that produce and release these molecules themselves. Release of TNFα from cultured synoviocytes from arthritic animals is reduced by application of CBD (Malfait et al., 2000) as well as by TRPV1 antagonists (Kochukov et al., 2009). Desensitization of TRPV1 by CBD could thus interrupt further progression of this cycle. Further studies are needed to identify specific receptors and mechanisms underlying the anti-inflammatory and anti-hyperalgesic effects of CBD.
The pro-inflammatory/pro-pain spiral is further driven by similar mechanisms which induce the expression of pro-inflammatory biomarkers in the DRG of the afferent nerves and centrally in the spinal cord. Transdermal CBD application was successful in decreasing monoarthritis-associated increases of pro-inflammatory biomarkers in neuronal tissues. Here, adjuvant-induced monoarthritis led to significant increases of TNFα expression in DRG which was decreased to baseline levels by transdermal CBD application.
In this study, the expression of CGRP in spinal cord was increased after peripheral inflammation as previously reported (Sluka and Westlund, 1993), and was decreased by high doses (6.2 and 62 mg/day) of transdermal CBD. Like other neuropeptides, CGRP is rapidly transported to nerve terminals for release centrally as well as peripherally where as a potent vasodilator it contributes to neurogenic inflammation (Kawasaki et al., 1988; Zhang et al., 2001). Although increases in CGRP are described in DRG after hindpaw inflammation (Nahin and Byers, 1994), no significant changes in DRG expression were observed here (data not shown), possibly due to the small number of sensory neurons innervating the joint. It is also plausible that by 7 days post monoarthritis induction, neuropeptide content has stabilized.
In monoarthritic animals, spinal cord OX42 expression is increased in activated microglia, the immune cells specific to the central nervous system, as previously described for CFA-induced ankle inflammation and trinitrobenzene sulfonic acid (TNBS)-induced pancreatitis (Shan et al., 2007; Liu et al., 2012). Treatment with high doses of transdermal CBD in this study effectively reduced OX42 expression below baseline levels, indicating reduced microglial activation. CBD also reduced microglial/macrophage spinal cord expression in a mouse encephalopathy model (Kozela et al., 2011).
4.4 Absence of psychoactive behaviours
CBD is a known non-psychoactive cannabinoid, and due to its low affinity for the CB1 receptor it would be expected that exploratory behavioural activity would be similar among treatment groups compared to negative side-effects associated with THC (Croxford, 2003; Malone et al., 2009). This was demonstrated in this study by lack of CBD-induced changes in open-field exploration among naive treatment groups. Combinatorial with psychoactivity, side-effects such as hypothermia and hypomobility induced by THC are avoided with use of CBD (Zimmer et al., 1999).
5. Conclusion
These studies demonstrate transdermal administration of CBD has long-lasting therapeutic effects without psychoactive side-effects. Thus, use of topical CBD has potential as effective treatment of arthritic symptomatology. At present, one in five (21%) adults worldwide are diagnosed with some form of arthritis by their physicians (Helmick et al., 2008). The data presented suggest transdermal CBD is a good candidate for developing improved therapies for these debilitating disease.
What’s already known about this topic?
- Cannabinoids and cannabinoid receptors have been studied as potential targets for reducing pain and inflammation associated with osteoarthritis and rheumatoid arthritis. Cannabinoid side-effects vary and depend on several factors like administrated dose, route of administration, etc.
- Transdermal cannabidiol (CBD) gel application has therapeutic potential for relief of arthritic pain-related behaviours and exerts an anti-inflammation property without evident high brain centre psychoactive effects.
Acknowledgments
Funding sources
This work was supported by the American Cancer Society award (RGS-00-027-04-CDD) to A.L. Stinchcomb, University of Kentucky start-up funds to K.N. Westlund and All-Tranz, Inc. Cannabidiol was a generous gift provided by NIDA.
Footnotes
Conflicts of interest
None declared.
Author contributions
D.C.H. prepared and applied the treatments, tested blood drug concentrations, performed data analysis and wrote portions of the first draft of this manuscript. L.Z. performed the drug application, behavioural tests, data analysis, statistics and produced the figures, as well as drafted portions of and edited this manuscript. F.M. performed the blinded behavioural tests, read and edited this manuscript. S.M.A. performed the blinded behavioural tests and read this manuscript. S.L.M. performed the immunostaining, data analysis, produced the figures, as well as read and edited this manuscript. A.L.S. designed the study, supplied the drug, read and edited this manuscript. K.N.W. designed the study, read and edited this manuscript. All authors discussed the results and commented on the manuscript.
References
- Akopian AN. Regulation of nociceptive transmission at the periphery via TRPA1-TRPV1 interactions. Curr Pharm Biotechnol. 2011;12:89–94. [PubMed]
- Akopian AN, Ruparel NB, Patwardhan A, Hargreaves KM. Cannabinoids desensitize capsaicin and mustard oil responses in sensory neurons via TRPA1 activation. J Neurosci. 2008;8:1064–1075.[PubMed]
- Akopian AN, Ruparel NB, Jeske NA, Patwardhan A, Hargreaves KM. Role of ionotropic cannabinoid receptors in peripheral antipain and antihyperalgesia. Trends Pharmacol Sci. 2009;30:79–84. [PMC free article] [PubMed]
- Begg M, Pacher P, Batkai S, Osei-Hyiaman D, Offertaler L, Mo FM, Liu J, Kunos G. Evidence for novel cannabinoid receptors. Pharmacol Ther. 2005;106:133–145. [PubMed]
- Berckmans RJ, Nieuwland R, Kraan MC, Schaap MC, Pots D, Smeets TJ, Sturk A, Tak PP. Synovial microparticles from arthritic patients modulate chemokine and cytokine release by synoviocytes. Arthritis Res Ther. 2005;7:R536–R544. [PMC free article] [PubMed]
- Berman JS, Symonds C, Birch R. Efficacy of two cannabis based medicinal extracts for relief of central neuropathic pain from brachial plexus avulsion: Results of a randomised controlled trial. Pain. 2004;112:299–306. [PubMed]
- Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol. 2001;134:845–852. [PMC free article] [PubMed]
- Brown AJ. Novel cannabinoid receptors. Br J Pharmacol. 2007;152:567–575. [PMC free article][PubMed]
- Burstein SH, Zurier RB. Cannabinoids, endocannabinoids, and related analogs in inflammation. AAPS J. 2009;11:109–119. [PMC free article] [PubMed]
- Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D. Impaired pain and pain sensation in mice lacking the capsaicin receptor. Science. 2000;288:306–313. [PubMed]
- Centers for Disease Control and Prevention (CDC) Prevalence of doctor-diagnosed arthritis and arthritis-attributable activity limitation – United States, 2007–2009. MMWR Morb Mortal Wkly Rep. 2010;59:1261–1265. [PubMed]
- Clayton N, Marshall FH, Bountra C, O’Shaughnessy CT. CB1 and CB2 cannabinoid receptors are implicated in inflammatory pain. Pain. 2002;96:253–260. [PubMed]
- Coffey RG, Yamamoto Y, Snella E, Pross S. Tetrahydrocannabinol inhibition of macrophage nitric oxide production. Biochem Pharmacol. 1996;52:743–751. [PubMed]
- Costa B, Colleoni M, Conti S, Parolaro D, Franke C, Trovato AE, Giagnoni G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn Schmiedebergs Arch Pharmacol. 2004a;369:294–299. [PubMed]
- Costa B, Giagnoni G, Franke C, Trovato AE, Colleoni M. Vanilloid TRPV1 receptor mediates the antihyperalgesic effect of the nonpsychoactive cannabinoid, cannabidiol, in a rat model of acute inflammation. Br J Pharmacol. 2004b;143:247–250. [PMC free article] [PubMed]
- Costa B, Trovato AE, Comelli F, Giagnoni G, Colleoni M. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur J Pharmacol. 2007;556:75–83. [PubMed]
- Crawford M, Curtis JR. Tumor necrosis factor inhibitors and infection complications. Curr Rheumatol Rep. 2008;10:383–389. [PMC free article] [PubMed]
- Croxford JL. Therapeutic potential of cannabinoids in CNS disease. CNS Drugs. 2003;17:179–202.[PubMed]
- Cunnane G, Bjork L, Ulfgren AK, Lindblad S, FitzGerald O, Bresnihan B, Andersson U. Quantitative analysis of synovial membrane inflammation: A comparison between automated and conventional microscopic measurements. Ann Rheum Dis. 1999;58:493–499. [PMC free article][PubMed]
- Davis JB, Gray J, Gunthorpe MJ, Hatcher JP, Davey PT, Overend P, Harries MH, Latcham J, Clapham C, Atkinson K, Hughes SA, Rance K, Grau E, Harper AJ, Pugh PL, Rogers DC, Bingham S, Randall A, Sheardown SA. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature. 2000;405:183–187. [PubMed]
- Fioravanti B, De Felice M, Stucky CL, Medler KA, Luo MC, Gardell LR, Ibrahim M, Malan TP, Jr, Yamamura HI, Ossipov MH, King T, Lai J, Porreca F, Vanderah TW. Constitutive activity at the cannabinoid CB1 receptor is required for behavioral response to noxious chemical stimulation of TRPV1: Antinociceptive actions of CB1 inverse agonists. J Neurosci. 2008;28:11593–11602.[PMC free article] [PubMed]
- Furst DE. Development of TNF inhibitor therapies for the treatment of rheumatoid arthritis. Clin Exp Rheumatol. 2010;28:S5–S12. [PubMed]
- Godlewski G, Offertaler L, Wagner JA, Kunos G. Receptors for acylethanolamides-GPR55 and GPR119. Prostaglandins Other Lipid Mediat. 2009;89:105–111. [PMC free article] [PubMed]
- Guerne PA, Terkeltaub R, Zuraw B, Lotz M. Inflammatory microcrystals stimulate interleukin-6 production and secretion by human monocytes and synoviocytes. Arthritis Rheum. 1989;32:1443–1452. [PubMed]
- Guo LH, Schluesener HJ. Acute but not chronic stimulation of glial cells in rat spinal cord by systemic injection of lipopolysaccharide is associated with hyperalgesia. Acta Neuropathol. 2006;112:703–713. [PubMed]
- Hastings R, Ding T, Butt S, Gadsby K, Zhang W, Moots RJ, Deighton C. Neutropenia in patients receiving anti-tumor necrosis factor therapy. Arthritis Care Res (Hoboken) 2010;62:764–769.[PubMed]
- Helmick CG, Felson DT, Lawrence RC, Gabriel S, Hirsch R, Kwoh CK, Liang MH, Kremers HM, Mayes MD, Merkel PA, Pillemer SR, Reveille JD, Stone JH. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States Part I. Arthritis Rheum. 2008;58:15–25.[PubMed]
- Kawasaki H, Takasaki K, Saito A, Goto K. Calcitonin gene-related peptide acts as a novel vasodilator neurotransmitter in mesenteric resistance vessels of the rat. Nature. 1988;335:164–167.[PubMed]
- Keeble J, Russell F, Curtis B, Starr A, Pinter E, Brain SD. Involvement of transient receptor potential vanilloid 1 in the vascular and hyperalgesic components of joint inflammation. Arthritis Rheum. 2005;52:3248–3256. [PubMed]
- Kerstein PC, del Camino D, Moran MM, Stucky CL. Pharmacological blockade of TRPA1 inhibits mechanical firing in nociceptors. Mol Pain. 2009;5:19. [PMC free article] [PubMed]
- Kochukov MY, McNearney TA, Yin H, Zhang L, Ma F, Ponomareva L, Abshire S, Westlund KN. Tumor necrosis factor-alpha (TNF-alpha) enhances functional thermal and chemical responses of TRP cation channels in human synoviocytes. Mol Pain. 2009;5:49. [PMC free article] [PubMed]
- Kozela E, Lev N, Kaushansky N, Eilam R, Rimmerman N, Levy R, Ben-Nun A, Juknat A, Vogel Z. Cannabidiol inhibits pathogenic T cells, decreases spinal microglial activation and ameliorates multiple sclerosis-like disease in C57BL/6 mice. Br J Pharmacol. 2011;163:1507–1519.[PMC free article] [PubMed]
- Kress M, Kuner R. Mode of action of cannabinoids on nociceptive nerve endings. Exp Brain Res. 2009;196:79–88. [PubMed]
- Liu PY, Lu CL, Wang CC, Lee IH, Hsieh JC, Chen CC, Lee HF, Lin HC, Chang FY, Lee SD. Spinal microglia initiate and maintain hyperalgesia in a rat model of chronic pancreatitis. Gastroenterology. 2012;142:165–173. e162. [PubMed]
- Lodzki M, Godin B, Rakou L, Mechoulam R, Gallily R, Touitou E. Cannabidiol-transdermal delivery and anti-inflammatory effect in a murine model. J Control Release. 2003;93:377–387.[PubMed]
- Malfait AM, Gallily R, Sumariwalla PF, Malik AS, Andreakos E, Mechoulam R, Feldmann M. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc Natl Acad Sci USA. 2000;97:9561–9566. [PMC free article][PubMed]
- Malone DT, Jongejan D, Taylor DA. Cannabidiol reverses the reduction in social interaction produced by low dose Delta(9)- tetrahydrocannabinol in rats. Pharmacol Biochem Behav. 2009;93:91–96. [PubMed]
- Malone DT, Hill MN, Rubino T. Adolescent cannabis use and psychosis: Epidemiology and neurodevelopmental models. Br J Pharmacol. 2010;160:511–522. [PMC free article] [PubMed]
- McGaraughty S, Chu KL, Perner RJ, Didomenico S, Kort ME, Kym PR. TRPA1 modulation of spontaneous and mechanically evoked firing of spinal neurons in uninjured, osteoarthritic, and inflamed rats. Mol Pain. 2010;6:14. [PMC free article] [PubMed]
- Mechoulam R. Marihuana chemistry. Science. 1970;168:1159–1166. [PubMed]
- Mechoulam R, Hanus L. Cannabidiol: An overview of some chemical and pharmacological aspects. Part I: Chemical aspects. Chem Phys Lipids. 2002;121:35–43. [PubMed]
- Mechoulam R, Shvo Y. Hashish. 1. Structure of cannabidiol. Tetrahedron. 1963;19:2073–2074.[PubMed]
- Mechoulam R, Parker LA, Gallily R. Cannabidiol: An overview of some pharmacological aspects. J Clin Pharmacol. 2002;42:11S–19S. [PubMed]
- Nahin RL, Byers MR. Adjuvant-induced inflammation of rat paw is associated with altered calcitonin gene-related peptide immunoreactivity within cell bodies and peripheral endings of primary afferent neurons. J Comp Neurol. 1994;349:475–485. [PubMed]
- Neumann E, Lefevre S, Zimmermann B, Gay S, Muller-Ladner U. Rheumatoid arthritis progression mediated by activated synovial fibroblasts. Trends Mol Med. 2010;16:458–468. [PubMed]
- Ospelt C, Neidhart M, Gay RE, Gay S. Synovial activation in rheumatoid arthritis. Front Biosci. 2004;9:2323–2334. [PubMed]
- Paudel KS, Milewski M, Swadley CL, Brogden NK, Ghosh P, Stinchcomb AL. Challenges and opportunities in dermal/transdermal delivery. Ther Deliv. 2010;1:109–131. [PMC free article][PubMed]
- Petrus M, Peier AM, Bandell M, Hwang SW, Huynh T, Olney N, Jegla T, Patapoutian A. A role of TRPA1 in mechanical hyperalgesia is revealed by pharmacological inhibition. Mol Pain. 2007;3:40.[PMC free article] [PubMed]
- Richardson D, Pearson RG, Kurian N, Latif ML, Garle MJ, Barrett DA, Kendall DA, Scammell BE, Reeve AJ, Chapman V. Characterisation of the cannabinoid receptor system in synovial tissue and fluid in patients with osteoarthritis and rheumatoid arthritis. Arthritis Res Ther. 2008;10:R43.[PMC free article] [PubMed]
- Roberts J, Ossipov MH, Porreca F. Glial activation in the rostroventromedial medulla promotes descending facilitation to mediate inflammatory hypersensitivity. Eur J Neurosci. 2009;30:229–241.[PMC free article] [PubMed]
- Ryberg E, Larsson N, Sjogren S, Hjorth S, Hermansson NO, Leonova J, Elebring T, Nilsson K, Drmota T, Greasley PJ. The orphan receptor GPR55 is a novel cannabinoid receptor. Br J Pharmacol. 2007;152:1092–1101. [PMC free article] [PubMed]
- Schafers M, Sorkin L. Effect of cytokines on neuronal excitability. Neurosci Lett. 2008;437:188–193. [PubMed]
- Shan S, Qi-Liang MY, Hong C, Tingting L, Mei H, Haili P, Yan-Qing W, Zhi-Qi Z, Yu-Qiu Z. Is functional state of spinal microglia involved in the anti-allodynic and anti-hyperalgesic effects of electroacupuncture in rat model of monoarthritis? Neurobiol Dis. 2007;26:558–568.[PMC free article] [PubMed]
- Sluka KA, Westlund KN. Behavioral and immunohistochemical changes in an experimental arthritis model in rats. Pain. 1993;55:367–377. [PubMed]
- Sluka KA, Willis WD, Westlund KN. Joint inflammation and hyperalgesia are reduced by spinal bicuculline. NeuroReport. 1993;5:109–112. [PubMed]
- Sluka KA, Lawand NB, Westlund KN. Joint inflammation is reduced by dorsal rhizotomy and not by sympathectomy or spinal cord transection. Ann Rheum Dis. 1994;53:309–314. [PMC free article][PubMed]
- Sorkin LS, Doom CM. Epineurial application of TNF elicits an acute mechanical hyperalgesia in the awake rat. J Peripher Nerv Syst. 2000;5:96–100. [PubMed]
- Staton PC, Hatcher JP, Walker DJ, Morrison AD, Shapland EM, Hughes JP, Chong E, Mander PK, Green PJ, Billinton A, Fulleylove M, Lancaster HC, Smith JC, Bailey LT, Wise A, Brown AJ, Richardson JC, Chessell IP. The putative cannabinoid receptor GPR55 plays a role in mechanical hyperalgesia associated with inflammatory and neuropathic pain. Pain. 2008;139:225–236. [PubMed]
- Stinchcomb AL, Valiveti S, Hammell DC, Ramsey DR. Human skin permeation of Delta8-tetrahydrocannabinol, cannabidiol and cannabinol. J Pharm Pharmacol. 2004;56:291–297. [PubMed]
- Szabo A, Helyes Z, Sandor K, Bite A, Pinter E, Nemeth J, Banvolgyi A, Bolcskei K, Elekes K, Szolcsanyi J. Role of transient receptor potential vanilloid 1 receptors in adjuvant-induced chronic arthritis: In vivo study using gene-deficient mice. J Pharmacol Exp Ther. 2005;314:111–119.[PubMed]
- Turner CE, Bouwsma OJ, Billets S, Elsohly MA. Constituents of Cannabis sativa L. XVIII–Electron voltage selected ion monitoring study of cannabinoids. Biomed Mass Spectrom. 1980;7:247–256.[PubMed]
- Westlund KN, Kochukov MY, Lu Y, McNearney TA. Impact of central and peripheral TRPV1 and ROS levels on proinflammatory mediators and nociceptive behavior. Mol Pain. 2010;6:46.[PMC free article] [PubMed]
- Whyte LS, Ryberg E, Sims NA, Ridge SA, Mackie K, Greasley PJ, Ross RA, Rogers MJ. The putative cannabinoid receptor GPR55 affects osteoclast function in vitro and bone mass in vivo. Proc Natl Acad Sci USA. 2009;106:16511–16516. [PMC free article] [PubMed]
- Zhang L, Hoff AO, Wimalawansa SJ, Cote GJ, Gagel RF, Westlund KN. Arthritic calcitonin/alpha calcitonin gene-related peptide knockout mice have reduced nociceptive hypersensitivity. Pain. 2001;89:265–273. [PubMed]
- Zhang L, Lu Y, Chen Y, Westlund KN. Group I metabotropic glutamate receptor antagonists block secondary thermal hyperalgesia in rats with knee joint inflammation. J Pharmacol Exp Ther. 2002;300:149–156. [PubMed]
- Zhang L, Zhang X, Westlund KN. Restoration of spontaneous exploratory behaviors with an intrathecal NMDA receptor antagonist or a PKC inhibitor in rats with acute pancreatitis. Pharmacol Biochem Behav. 2004;77:145–153. [PMC free article] [PubMed]
- Zimmer A, Zimmer AM, Hohmann AG, Herkenham M, Bonner TI. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc Natl Acad Sci USA. 1999;96:5780–5785. [PMC free article] [PubMed]
- Zuardi AW. Cannabidiol: From an inactive cannabinoid to a drug with wide spectrum of action. Rev Bras Psiquiatr. 2008;30:271–280. [PubMed]
Last edited: